Even though the topic is called animal physiology, most of the content is directed towards human physiology. The topic explores concepts already covered in topic 6 with more depth, in addition to introducing new concepts. I liked the topic very much and I did not find it harder than topic 6 at all. This topic completes the investigation into most of the major human organs with the notable exception of the brain and liver. I think it is a good way to end off the mandatory higher level curriculum.
Points for revision:
- The immune system in mammals
- Nitrogenous wastes in different organisms
- Fertilization in other animals
- Early development of a fetus
The immune system in mammals
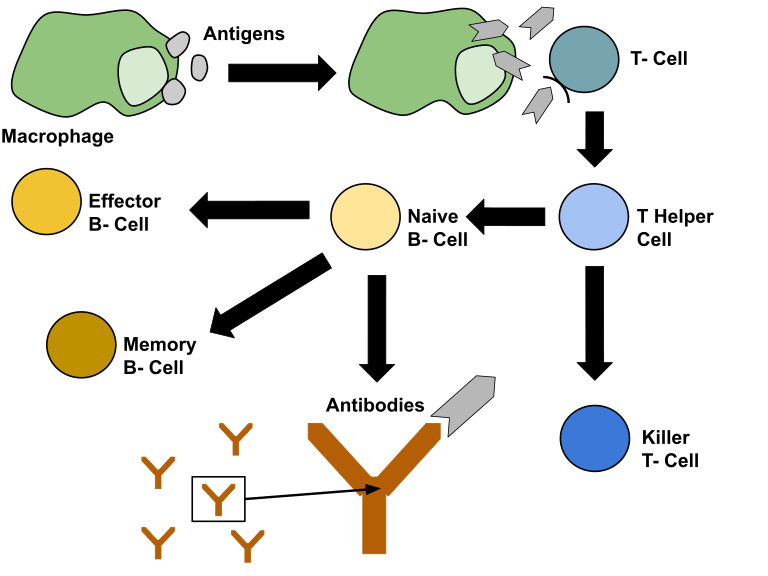
When the first layer of defense (the skin) is breached, the body initiates its second layer of defense, the immune system. The immune system is initiated when leucocytes (plasma cells) recognize cells in the body as “non self”. This is done by recognizing the surface proteins of cells. If the leucocytes detect a cell that does not correspond with the body cells, it sounds an alarm signaling the initiation of the immune system. The first cell involved in the immune response is the large macrophage. This cell can devour intruding pathogens and partially digest it chemically. It then presents parts of the pathogen on its surface in a process called antigen presentation. Helper T cells that match with the antigen presented then communicate with helper B cells that also match with the antigen. The immune response then turns from a non-specific response into an antigen-specific response. The activated B cells clones itself by mitosis, producing memory cells in the same process. Memory cells are long lived and provide the body with specific immunity in the case of a secondary infection. The activated B cells work by producing antibodies. Antibodies are small proteins that attach to the surface proteins of a specific pathogen, disabling them or marking them for other cells to destroy. The process of finding B cells that can produce the required antibodies is a time-consuming process, so the pathogens during a first-time encounter (primary immune response) usually have enough time to produce the symptoms related with the pathogen, thus making the host sick. True immunity is first acquired after the primary response by long lived memory cells floating around in the blood. When the same pathogen attacks, the secondary response is activated. During the secondary response, the body can immediately recognize the pathogen and tell the memory cells to start producing antibodies. The secondary response is usually so fast that no symptoms are developed. Vaccines work by exposing the immune system to disabled pathogens so that it can produce memory cells in the case of an actual infection. These memory cells can protect the body for a long time, but they will eventually die. When the concentration of memory cells is too low, a new dose of vaccine is needed in order to protect the body again.
{kind=link}
Extracting antibodies
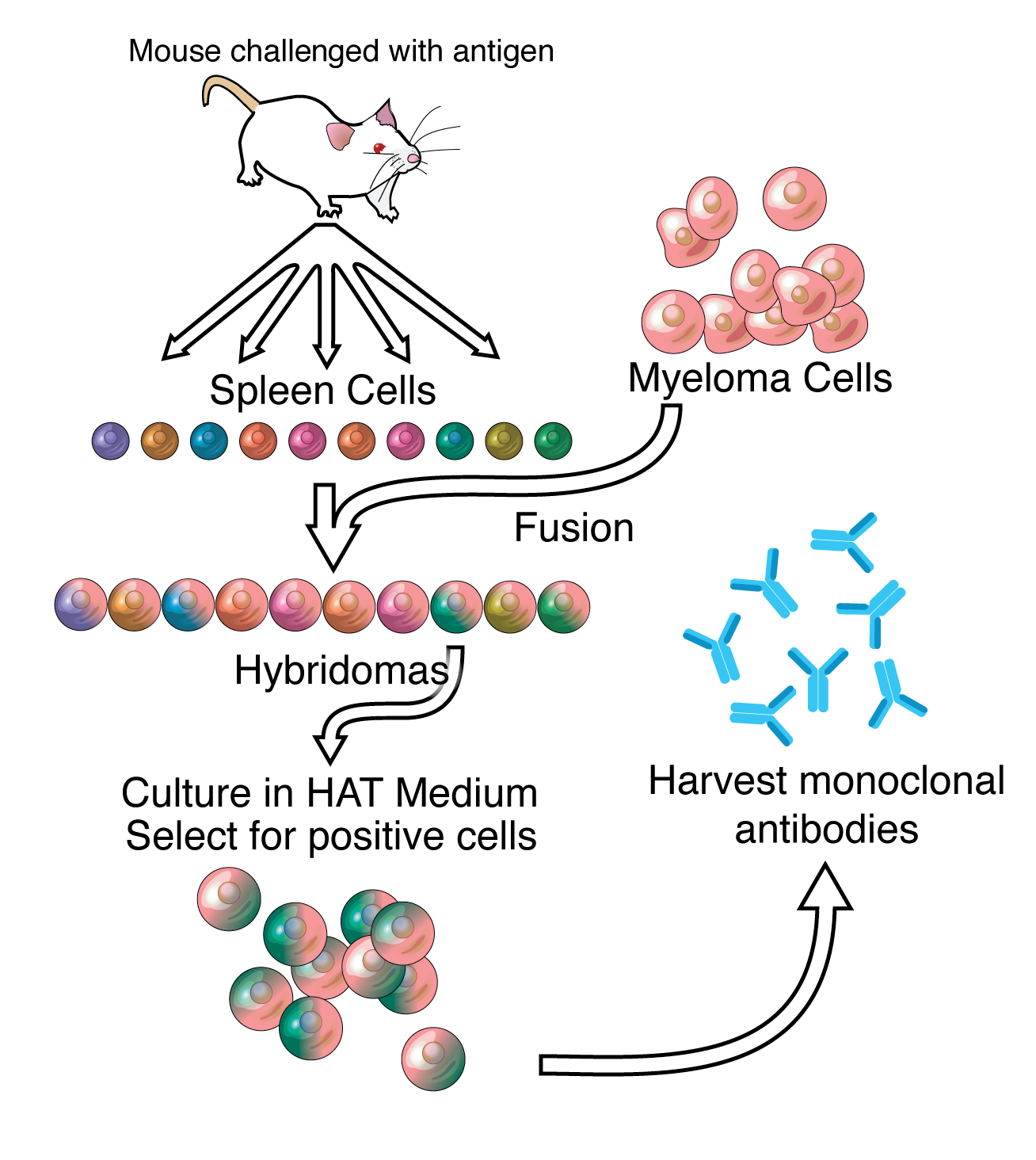
Antibodies can be extracted through fusing white blood cells with cancerous cells to make hybridoma cells. Hybridoma cells can produce antibodies because of the white blood cells, and they are long lived because of the cancerous cell. The process of extracting the desired antibody starts by infecting a host (for example a mouse) with a chose antigen to let the host carry out a primary immune response. During a primary response, multiple proteins are recognized as antigens, thus producing multiple types of antibodies. This is why a primary response is called polyclonal. In order to extract the desired antibody, multiple types of white blood cells are fused into hybridoma cells and tested by a protocol called ELISA (enzyme-linked immunosorbent assay). The white blood cells purely producing the desired antibodies are extracted and called monoclonal antibodies.
{kind=link}
Allergic reactions
Allergic reactions are a result of the body recognizing non-harmful (non-pathogenic) substances such as pollen as harmful (pathogenic) substances. Substances that can trigger an allergic reaction are called allergens. When the body detects allergens for the first time, the immune system is initiated to protect the body from the “invaders”. B cells start to produce antibodies called IgE antibodies which binds to mast cells containing vesicles filled with histamine. When the body is exposed to the allergen again, the allergen binds with the antibodies bonded to the mast cells, which causes the mast cells to release the vesicles filled with histamine. The release of histamine is associated with the symptoms of allergic reactions.
Skeletal muscle structure
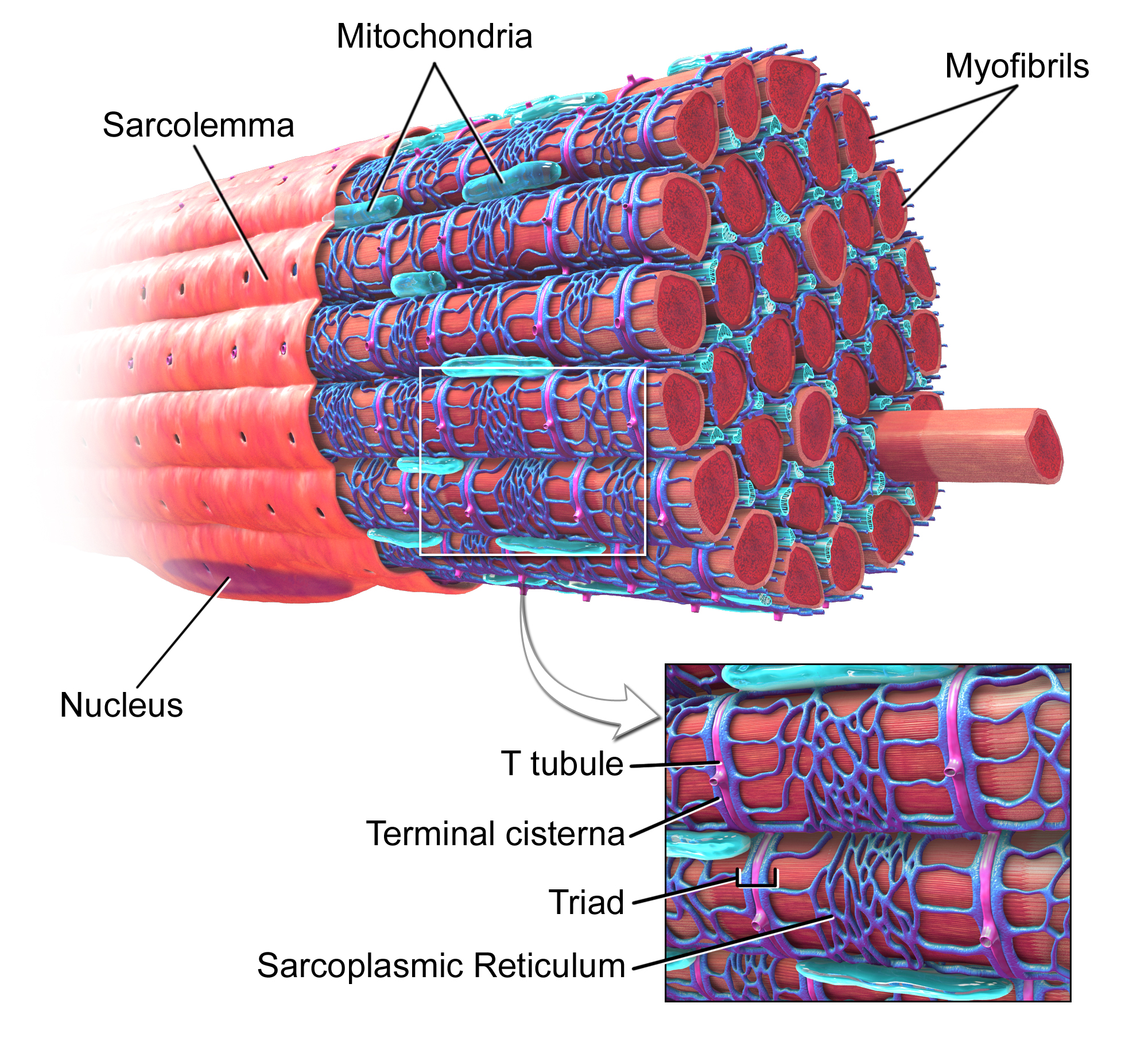
Skeletal muscles are made of bundles of muscle cells specialized for contracting and relaxing. Muscles cells are rich in mitochondria and have multiple nuclei. The cells themselves are made of a bundle of fibers called myofibrils which are inside a membrane called the sarcolemma. The sarcolemma has tube-like structures called transverse tubules (T tubules) that penetrates inside of the sarcolemma. The cytoplasm of the sarcolemma is called the sarcoplasm. Inside the sarcoplasm are all the organelles of the muscle cells, including organelles specialized for glycogen storage and oxygen storage. The organelle storing oxygen is called myoglobin. Skeletal muscles are connected to the skeleton or exoskeleton by tendons and pulls on the skeleton by contraction. Contraction happens inside the myofibrils by a set of two filaments called actin and myosin. Actin and myosin make up the contracting units of the muscle cell called sarcomeres, which are divided into sections by Z-lines. Myosin has “heads” which wants to attach to the myosin binding sites on the actin, but the actin is covered in two proteins called troponin and tropomyosin. Troponin and tropomyosin prevent the myosin from binding to the actin unless calcium ions are present.
{kind=link}
Muscle contraction
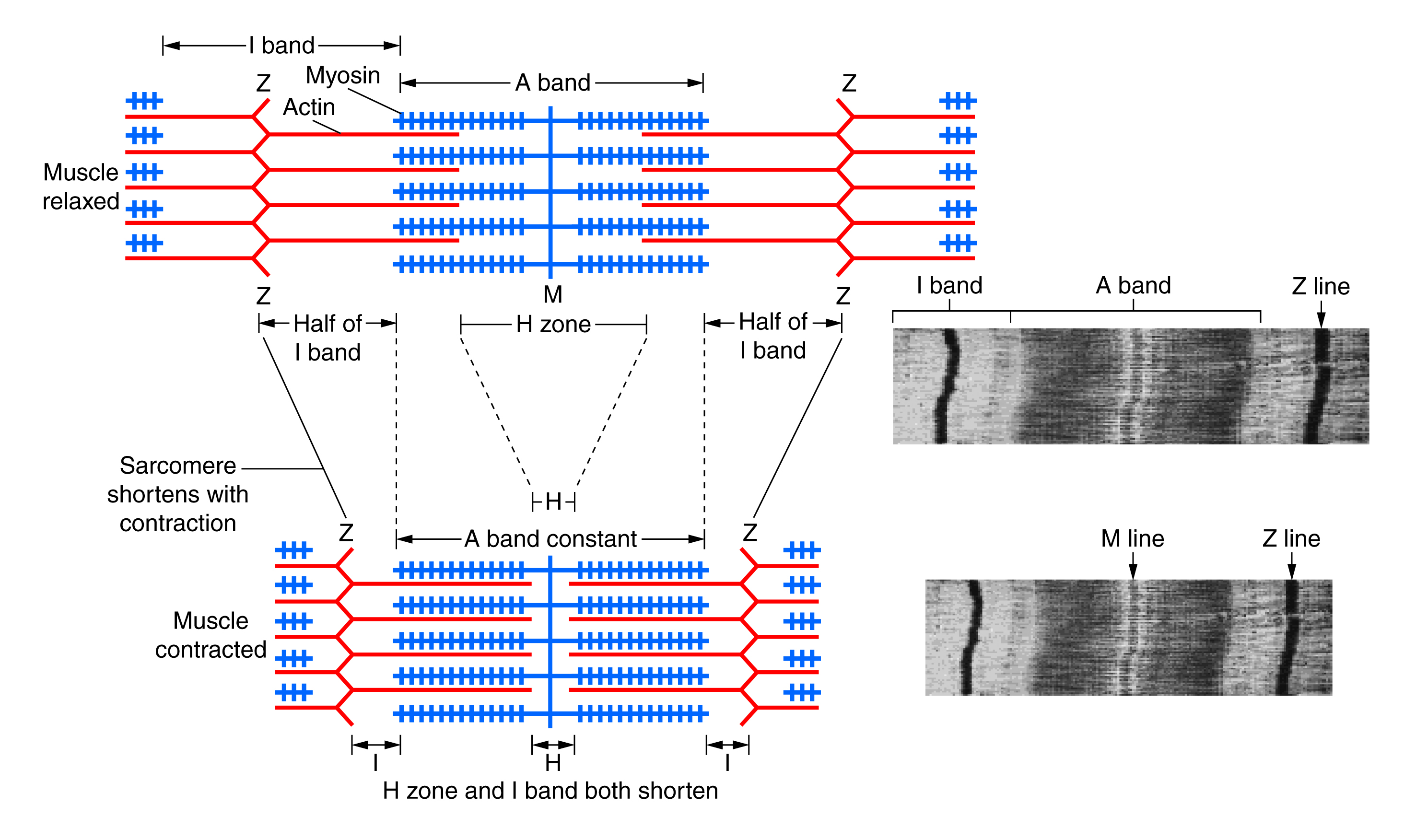
Contraction is the process of converting chemical energy into mechanical energy by muscle cells. When the muscle is not contracted, proteins called troponin and tropomyosin prevents the myosin from attaching to the actin by covering up the binding sites. Tropomyosin is a thin layer of proteins attached to troponin, which has binding sites for calcium. When the brain sends a signal down a motor neuron, voltage gated calcium channels in the T tubules inside the sarcoplasm opens, causing calcium ions to flow in. These calcium ions bind to the binding site on the troponin, which changes the shape of troponin. The change of shape in troponin pulls the tropomyosin away from the actin, thus exposing the binding sites for myosin. The myosin heads are already stretched and primed beforehand by the energy released by breaking up an ATP molecule into ADP and a phosphate group. When the binding sites on actin are exposed, the myosin heads are suddenly released like a spring, attaching to the actin pulling it towards the middle between the Z-lines. This motion causes the whole sarcomere to contract, thus contracting the muscle. After the ADP and phosphate are spent, they detach from the myosin head which causes a new ATP to bind with it. This binding releases the myosin head from the actin, thus relaxing the muscle. The ATP then splits into ADP and phosphate again, priming the muscle for the next contraction.
Movement
Muscles in animals are attached to a skeleton or an exoskeleton, and when the muscle contracts, the skeleton moves with it. In animals with a skeleton such as a mouse, the muscles are attached on the outside of the skeleton. In animals with an exoskeleton such as a grasshopper, the muscles are attached on the inside. Muscles attached to two bones with a joint between them work in antagonistic pairs. One muscle bends the joint, while the other muscle stretches the joint. An example of antagonistic muscles is the biceps and triceps. The biceps bend the arm, while the triceps stretches the arm. When the muscle contracts, only one of the bones are supposed to move. For example, when contracting the quadriceps, only the tibia moves while the femur is static. The type of joint in the knee is called a synovial joint. Synovial joints only allow movement in one direction, and they connect two bones together. In between the two bones is a fluid called synovial fluid which reduces friction and absorbs compression together with cartilage. The synovial fluid also provides nutrients to the cartilage cells. Tendons connects bone to tissue, while ligaments connect bone to bone. Another type of joint is the type of joint in our shoulders, called a ball and socket joint. These joints have the same components as synovial joints but allows movement in two directions.
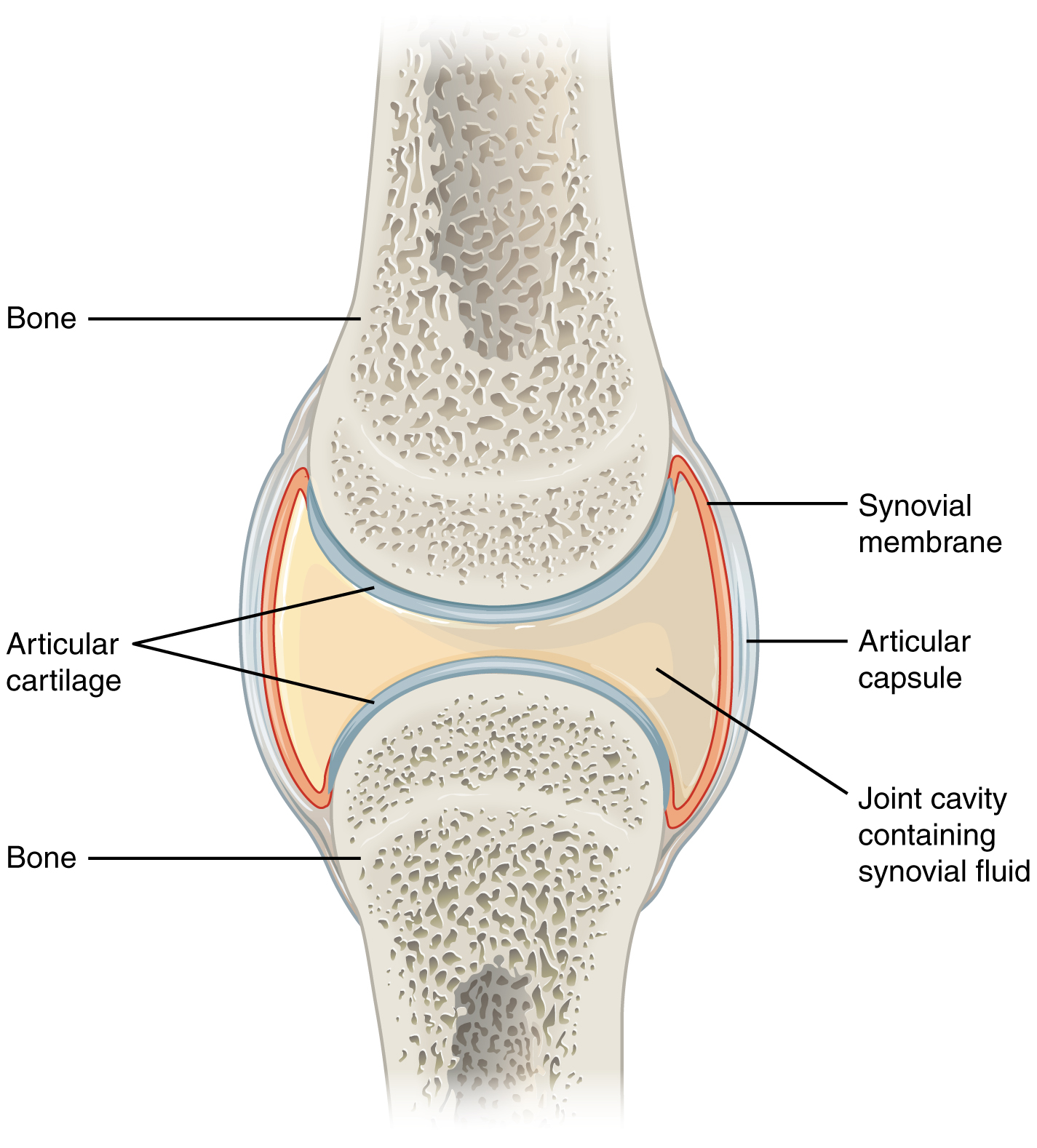
{kind=link}
Kidney and nephron anatomy
The kidneys function by filtrating waste products from the blood into urine, which is disposed through the urinal tract. The functional part of the kidneys are called nephrons, and the kidney is comprised of millions of them. Nephrons are composed of capillaries and a tubule, and functions by filtering out waste materials from the blood, and then reabsorbing the useful material which was also filtered out. The material leaving the nephrons are sent to the urine bladder. Humans have two kidneys that are connected to the blood system by the renal artery and the renal vein. Blood enters through the renal artery and is filtered in the nephrons. The filtered blood then leaves the kidneys through the renal vein. The inner part of the kidney is called the renal medulla, and the outer part is called the renal cortex. All the output of the nephrons are sent to the renal pelvis, which is connected to the urine bladder.
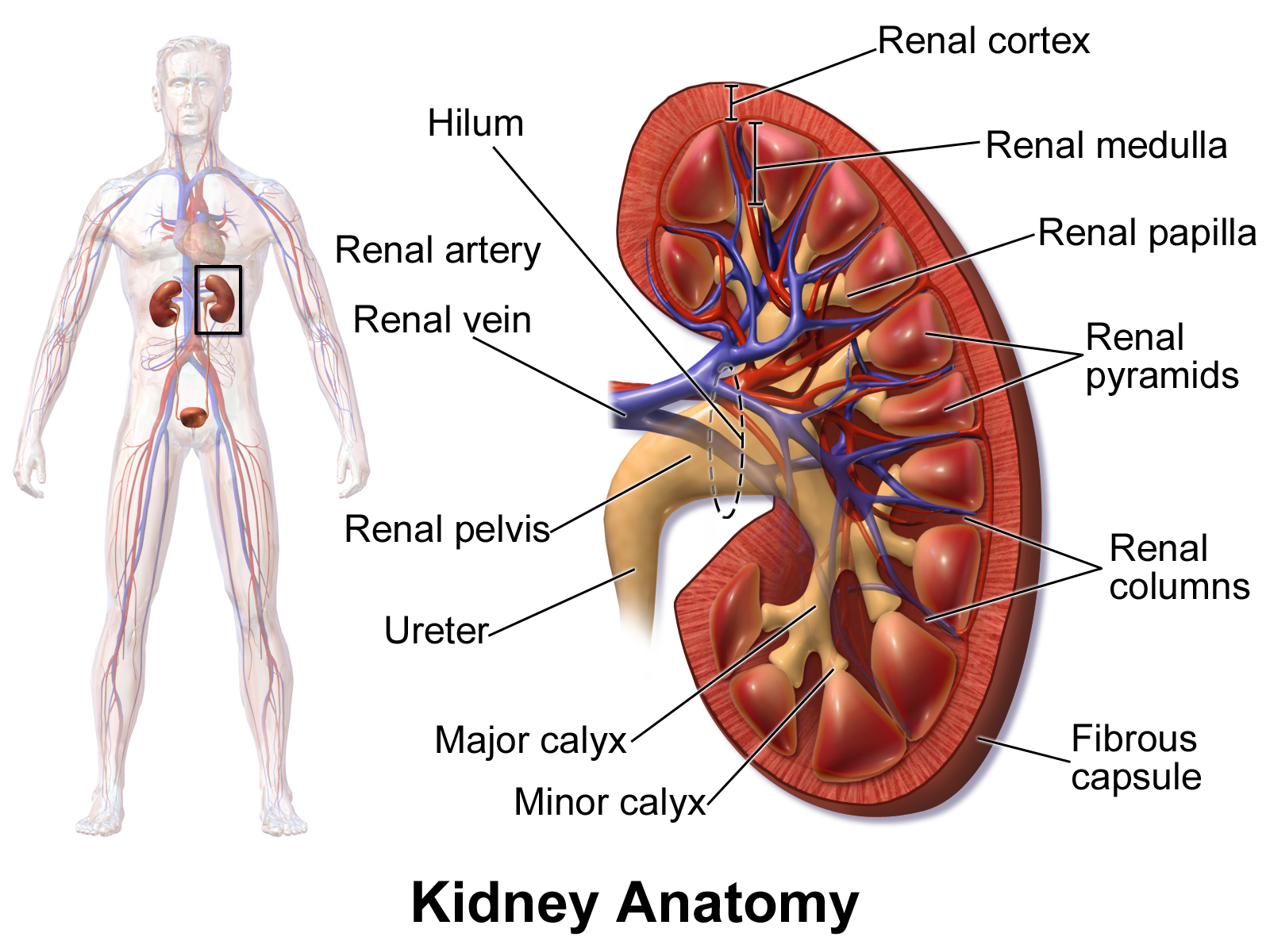
{kind=link}
Nephrons are comprised of capillaries and a long tubule. Unfiltered blood enters the nephrons from the afferent arterioles in the renal cortex (branches of the renal artery) and passes through a structure called Bowman’s capsule. Inside Bowman’s capsule, the capillaries bundle up into a capillary bed structure called the glomerulus. The high pressure inside the glomerulus filtrates blood out of the capillaries, and the filtrate is absorbed by Bowman’s capsule, entering the proximal convoluted tubule. The blood which is not filtrated in the glomerulus leaves Bowman’s capsule through the efferent arteriole and enters the peritubular capillary bed. The peritubular bed is important for reabsorption, and the blood that leaves the capillary bed is sent out into the body again through the renal vein. The filtrate in the proximal convoluted tubule enters a section of the tubule which dips down into the renal medulla called the loop of Henle. The part of the loop of Henle that takes the filtrate down into the medulla is permeable only to water, and the part that goes up into the renal cortex is permeable only to ions. After going up into the cortex again, the filtrate leaves the kidneys through the distal convoluted tubule and the collecting duct into the renal pelvis.
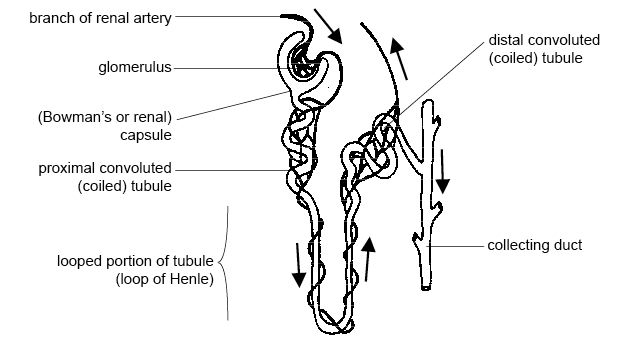
{kind=link}
Filtration
Filtration occurs as a result of the high pressure created when the afferent arteriole meets the glomerulus. The high pressure is built up by the blood vessels suddenly getting narrow, because when volume decreases, pressure increases. It is like a road that suddenly gets very narrow. Incoming cars have to slow down, thus increasing the concentration of cars in that area. Filtration occurs because the capillaries in the glomerulus are partially porous, which allows blood to be squeezed out of the capillary because of the high pressure. The ultrafiltered blood that is squeezed out of the glomerulus is then filtered through an additional basement membrane, which stops big molecules from leaving. The fluid leaving the basement membrane is called the filtrate and enters the proximal convoluted tubule. Despite all the filtering, there is still useful material in the filtrate which the body does not want to lose. The filtrate therefore must first go through a process of reabsorption before being disposed as urine.
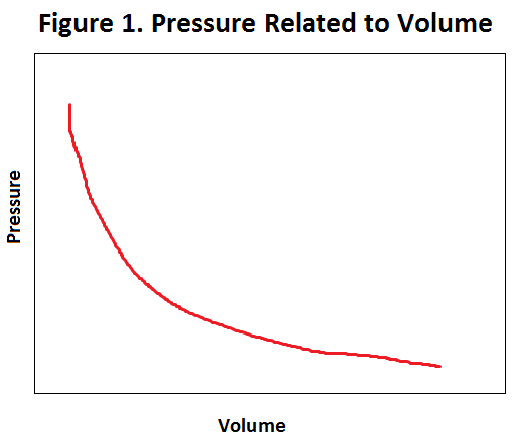
{kind=link}
Reabsorption
The filtrate filtrated by the glomerulus still contains valuable material, so the body has developed ways to get this material back by reabsorption. Reabsorption is made possible by the single layer thick convoluted tubule, collecting duct, and the nearby peritubular capillary bed. As previously mentioned, the ascending part of the loop of Henle in the medulla is permeable to ions. This is because it has sodium potassium pumps that pumps out ions by active transport, thus creating a hypertonic environment in the medulla. The hypertonic environment allows water to diffuse out of the convoluted tubule descending into the medulla by osmosis. In addition to ions and water being transported out, glucose is also pumped out by active transport. In healthy people urine should contain no glucose since it is all reabsorbed. The material that leaves the loop of Henle is reabsorbed by the peritubular capillary bed and recycled. Even though all of the valuable material is reabsorbed, the urine in the collecting duct is still too dilute. If some of the water in the urine would not be reabsorbed, the water intake would have to be increased drastically. This is why some of the water in the collecting duct is reabsorbed by water diffusing out as a result of semi permeability. The semi permeability of the colleting duct is dependent on the presence of a hormone produced by the posterior pituitary gland called ADH. When ADH is present, the collecting duct becomes more permeable to water, thus conserving more water. This is important for conserving water during times of drought and disposing water when it is in abundance. Animals such as desert animals have evolved longer tubes of Henle to be able to give it more surface area to diffuse water out, thus conserving water.

{kind=link}
Nitrogenous wastes in different organisms
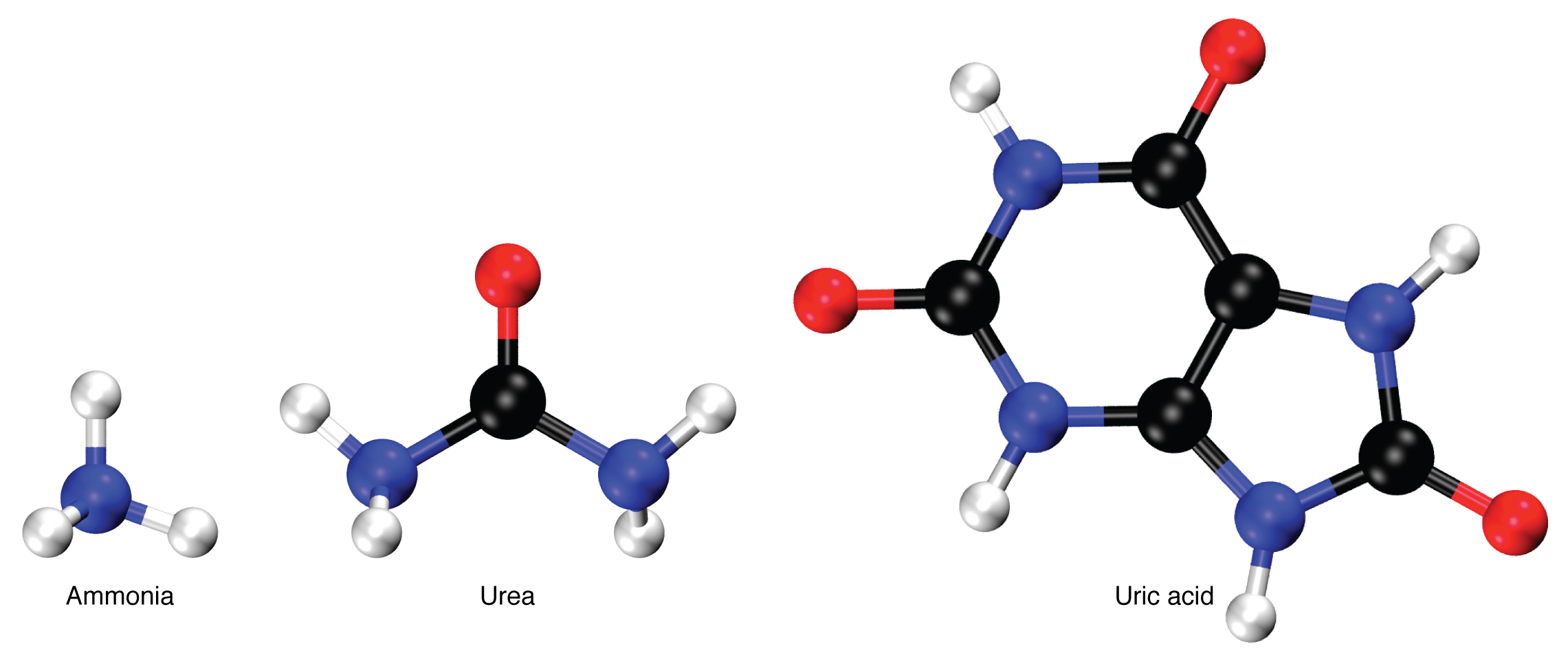
Since there is no good way to store unused amino acids, animals must dispose excess amino acids through nitrogenous wastes. Nitrogenous wastes get its name because they contain at least one nitrogen atom, which comes from the deamination of amino acids. Deamination is the process of removing the amino group (NH2) from amino acids. There are three types of nitrogenous wastes used by animals to dispose unused amino acids. The type of nitrogenous waste used highly depends on the environment and evolutionary past of the animal. Ammonia (NH3) is a simple molecule that does not require a lot of energy to produce. However, it is highly toxic, so it requires constant dilution and removal by water. Ammonia is usually used by animals living in aquatic environments such as fish. The abundance of water makes ammonia a suitable nitrogenous waste since it is cheap to produce, and easy to dispose. Urea (CH4N2O) is the nitrogenous waste used by humans and most mammals. It is more energy expensive to produce than ammonia, but it is less toxic, so it requires less water to dilute and remove. This makes it suitable for mammals which can produce abundant ATP and has water sources available. Uric acid (C5H4N4O3) is the most complex molecule, so it requires the most energy to produce. However, it is not harmful and can be easily stored. Furthermore, it requires little to no water to dispose. This makes uric acid a suitable nitrogenous waste for animals independent from aqueous environments such as birds.
{kind=link}
Spermatogenesis
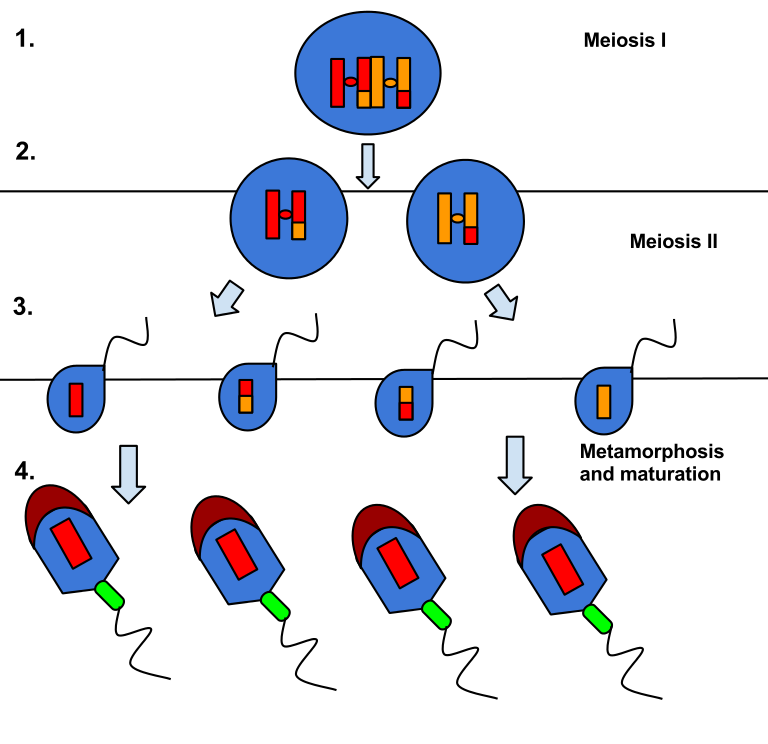
Sperm cells are the male reproductive cells and spermatogenesis is the process where they are created. Spermatogenesis occurs in the testes and is a process that starts during puberty and is continuous throughout the rest of the life cycle of a male. The testes themselves are located outside of the body to maintain a lower temperature than body temperature to ensure optimal environments. Inside the testes are numerous tubules called seminiferous tubules, and it is in these tubules where spermatogenesis occurs. Inside the seminiferous tubules are cells called spermatogonia which are reminiscent of stem cells in that they can reproduce by mitosis and “differentiate” by meiosis. The spermatogonia maintains its numbers by mitosis, then one of the replicated cells starts spermatogenesis by meiosis. The results of meiosis I and II are 4 haploid cells which can differentiate into sperm cells, also called spermatozoa. Differentiation happens in the seminiferous tubules when the haploid cells attach themselves to nutrient rich Sertoli cells located near the outer membrane of the seminiferous tubules. During differentiation, the cell grows a flagellum for mobility, and develops an acrosome containing the haploid nucleus. The acrosome is the head of the cell and is used during fertilization to reach inside of the ovule. The spermatozoa also develop a mitochondria storage to ensure sufficient energy for the journey. The composition of a spermatozoa is held minimal to be able to produce large amounts of cells. It seems like nature has selected quantity instead of quality.
{kind=link}
Oogenesis and development of an ovum
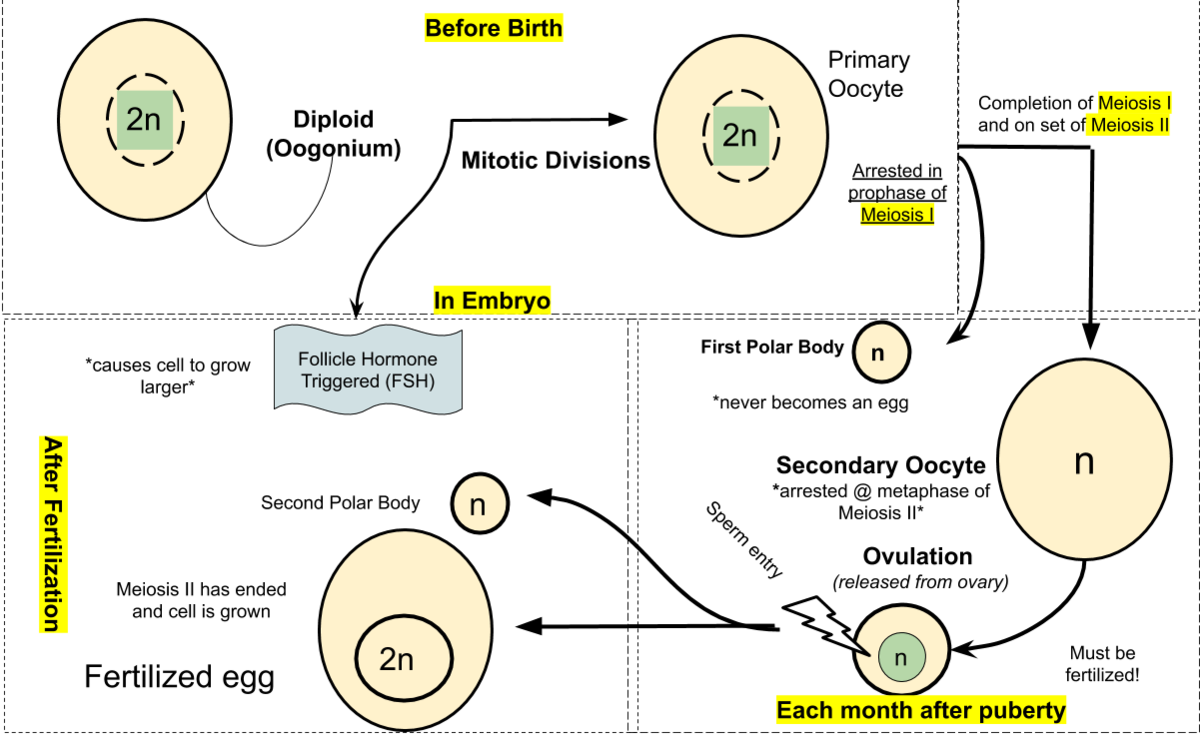
Female reproductive cells are called ova (plural of ovum) and oogenesis is the process where ova are created. Unlike spermatogenesis, all the ovum cells a female is ever going to have are developed before birth during the fetal stage. In the ovaries of a female fetus, diploid cells called oogonia starts to undergo mitosis, thus increasing their numbers. The oogonia cells then starts to grow into bigger cells called primary oocytes. Within the ovaries of females are cells called follicle cells, which develops simultaneously as oocytes and forms a protective ring around the primary oocyte. The entire structure is now called a primary follicle. The primary follicle starts meiosis but is arrested in prophase I until puberty. When a female reaches puberty, the primary oocytes start to mature into secondary oocytes, thus starting the menstrual cycle. Maturation of a primary oocyte into a secondary oocyte involves the cell finishing meiosis I, but unlike meiosis in spermatogenesis where all resulting cells are equal in size, only one ovum cell gains almost all the mass. Meiosis I result in two haploid cells where one cell is much larger than the other cell. The large cell goes on to developing into a secondary oocyte, while the smaller cell called a polar body undergoes cell death. After meiosis I, the follicle cells surrounding the oocyte undergoes mitosis, creating a second outer ring of follicle cells separated from the inner ring by a fluid filled cavity. This structure is called a graafian follicle, and as the cell grows, the pressure inside the fluid filled cavity starts to increase until the outer ring bursts and ovulation occurs. The secondary oocyte released in ovulation still has the inner follicle ring intact but has left the outer ring in the ovaries. The outer ring goes on to become the corpus luteum which releases progesterone and participates in regulating the menstrual cycle. The secondary oocyte has still not gone through meiosis II, and it does not do it until fertilization. If the oocyte is not fertilized, it undergoes cell death as the body prepares a new oocyte, starting the menstrual cycle over again.
{kind=link}
Fertilization in humans
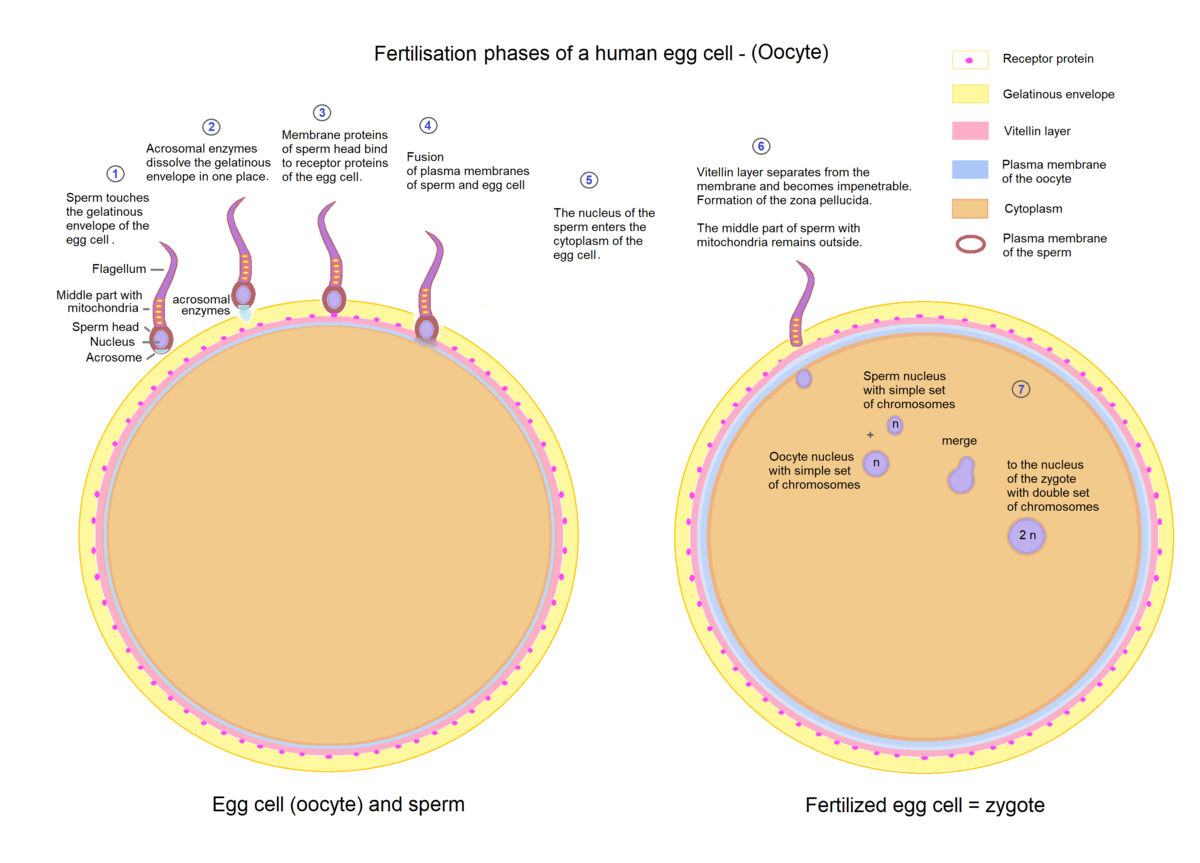
Fertilization is the process where a male sperm cell meets a female ovum and fuses into a zygote which eventually develops into a fetus. However, the sperm cell must first travel through the cervix and uterus before it can reach the ovum in the fallopian tube. Only a few sperm cells manage to make the journey. Furthermore, the sperm cells which successfully manages to reach the ovum, must then penetrate the follicle layer and a glycoprotein layer called the zona pellucida before reaching the oocyte. The zona pellucida is penetrated in a reaction called the acrosome reaction, where the sperm cell releases hydrolytic enzymes from its acrosome. The first sperm cell to reach the oocyte fuses its plasma membrane with the plasma membrane of the oocyte and releases its haploid nucleus into the cytoplasm. To prevent more than one sperm from cell fertilizing the ovum, a reaction called the cortical reaction is initiated. Granules in the cytoplasm of the oocyte called cortical granules fuses with the plasma membrane and releases enzymes that changes the chemical composition of the zona pellucida, making it impermeable for more sperm cells. Fertilization ends with the haploid nucleus of the sperm fusing with the haploid nucleus of the ovum, creating a diploid nucleus.
{kind=link}
Fertilization in other animals
Fertilization happens in a similar fashion in most animals, but there is a distinction between if the fertilization is internal or external. Most mammals use internal fertilization, where there is a strong maternal bond between the mother and newborn. It turns out that internal fertilization is the best way to ensure survival of the outcome. This is particularly valuable for big mammals who must spend a lot of energy in producing and developing the fetus. Other animals, such as fish, use external fertilization instead of internal fertilization. In external fertilization, the female lays its eggs outside of her body, awaiting a male to come by and fertilize them. External fertilization means a lower probability for a single egg to survive, but it also means that no energy is spent in developing the ovum. Animals who use external fertilization has solved the problem of low survival rates by laying large amounts of eggs. Even though the probability of a single egg surviving is quite low, the probability of at least one egg surviving is quite high.
Early development of a fetus
Shortly after fertilization has occurred, the fertilized ovum travels down the fallopian tube and implants itself in the highly vascular endometrium of the uterus. At this point the ovum has undergone a few mitotic divisions and become a ball of around 100 cells called a blastocyst. The blastocyst is surrounded by a layer of cells called the trophoblast and is around the same size as the ovum because the nutrients provided in the ovum has been used for metabolism instead of growth. The nutrients provided by the ovum are limited, so in order to keep a steady flow of nutrients to the ovum, the trophoblast layer and tissue from the uterus of the mother forms a placenta. The placenta further develops the umbilical cord which is comprised of three blood vessels for nutrient transport from the mother to fetus. Two of the blood vessels transports deoxygenized blood with waste materials to the placenta, and one blood vessel transports oxygenized and nutrient rich blood from the placenta. The blood of the mother and fetus never mixes because the blood vessels in the placenta are not connected. The placenta has capillary beds on the fetus side and on the mother side, so nutrients and other materials are exchanged back and forth by molecular transport.
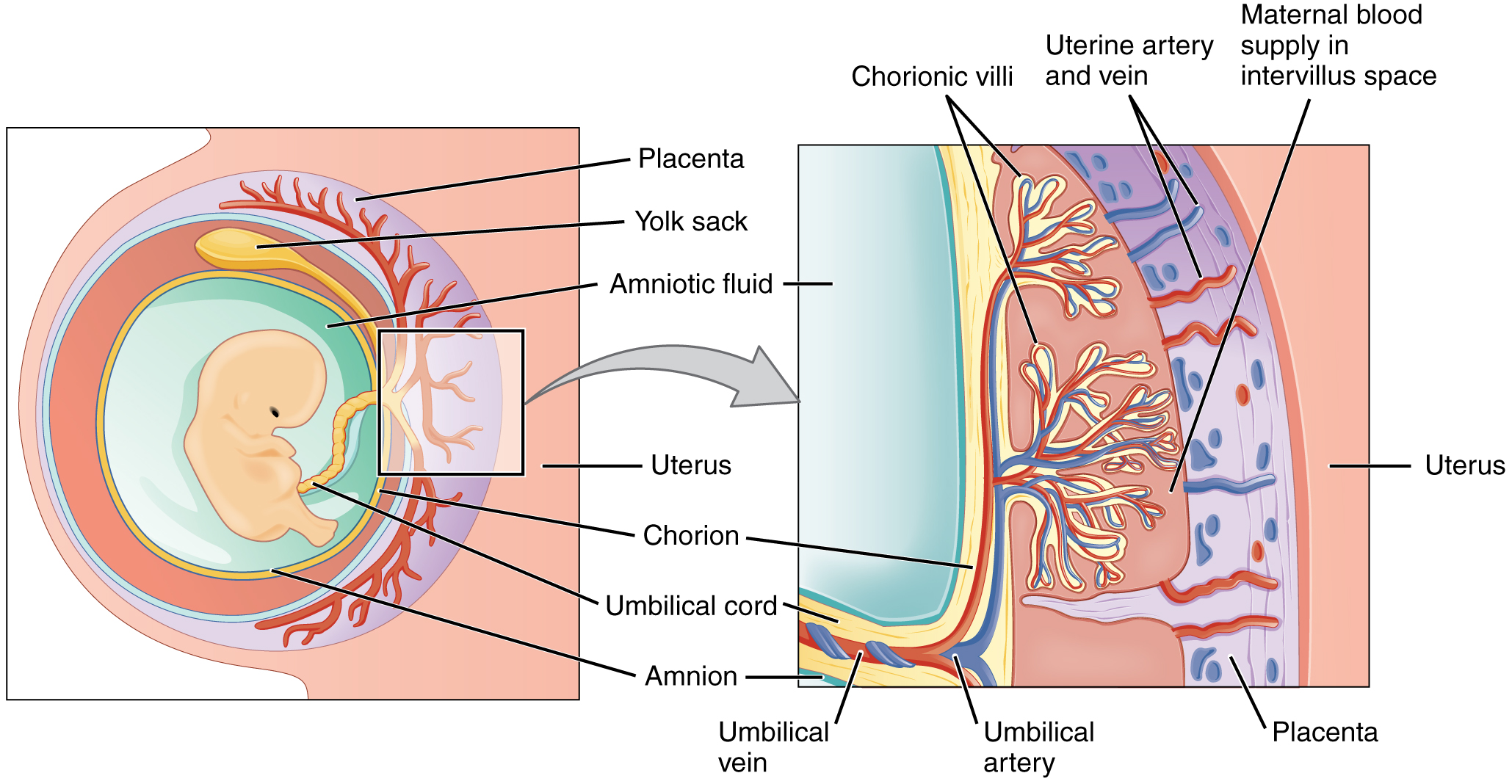
{kind=link}
Hormones during pregnancy
Usually, the corpus luteum within the ovaries of a non-pregnant female will degenerate a few days after ovulation, and with that the progesterone levels plummet and the highly vascular endometrium is released. However, when a female becomes pregnant the brain starts to secrete a hormone called human chorionic gonadotropin (HCG) which maintains the corpus luteum, thus maintaining progesterone levels and preserving the vascular endometrium for implantation to occur. After the placenta has developed, it takes over progesterone production in addition to starting production of high levels of estrogen. Progesterone maintains the highly vascular endometrium while estrogen prepares the female for childbirth. Both hormones prevent a new oocyte from developing, which is the basis for hormone regulating prevention pills. Another function of estrogen is to induce production of receptors in the uterine muscles which a hormone called oxytocin can bind to. Oxytocin is produced in the hypothalamus and released in the posterior pituitary by a positive feedback loop during pregnancy. When oxytocin binds to the receptors in the uterine muscles, the uterus contracts, which sends a signal to produce more oxytocin. Increased production of oxytocin means more frequent contractions which eventually culminates in childbirth.
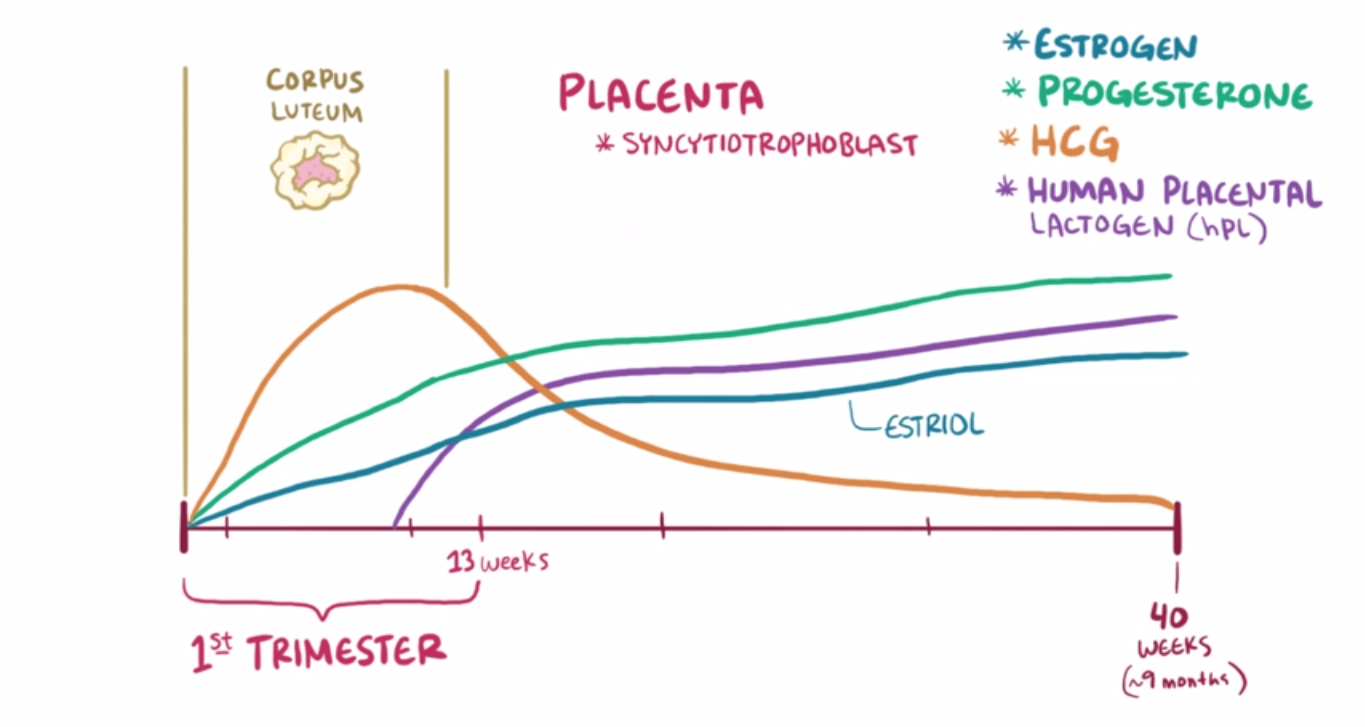
{kind=link}
Helpful videos:
Monoclonal antibodies: https://www.youtube.com/watch?v=kVs4VQQroOg
Skeletal muscles: https://www.youtube.com/watch?v=Ktv-CaOt6UQ
Kidneys and nephrons: https://www.youtube.com/watch?v=cc8sUv2SuaY
Spermatogenesis: https://www.youtube.com/watch?v=QUJ-xnv53UM
Oogenesis: https://www.youtube.com/watch?v=Msrym4oGVSk
Fertilization: https://www.youtube.com/watch?v=SUdAEGXLO-8
I very much enjoyed this topic even though I found topic 6 to be more interesting. I would say that the kidneys were the most interesting because I did not know a lot about them beforehand. None of the concepts were notably challenging, but it was a pretty lengthy topic with a few long and complicated words. As always, the diagrams in the book were very good, and there a plenty good Youtube videos available. Otherwise, I feel very satisfied with myself for finishing the curriculum (except one option).